Your new post is loading...
Your new post is loading...
Authors: Tian Su, Ziwei Li, Yinghua Zhang, Junqiang Xu and Bin Xu.
Journal of the American Society for Horticultural Science (2024)
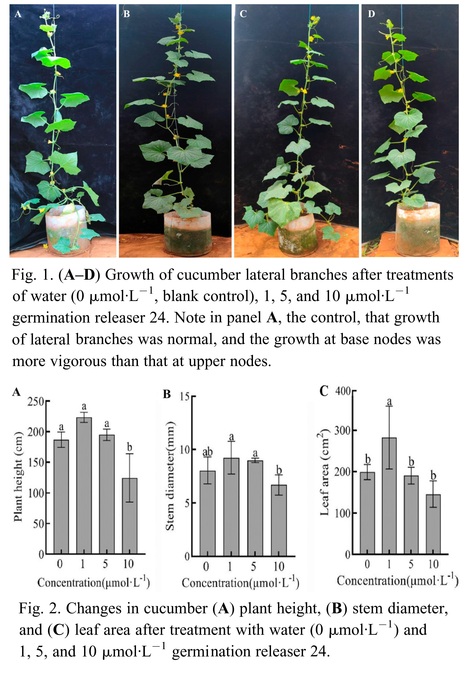
Abstract: "Cucumber (Cucumis sativus L.) belongs to the cucumber genus of the Cucurbitaceae family, and the selection of cultivars with minimal or no lateral branches can enhance the cultivation management efficiency. The growth of lateral branches is inhibited by strigolactone. To investigate the regulatory mechanism of strigolactone on the lateral branch development in cucumber, the cultivar LZ1 exhibiting multiple lateral branches was selected as the experimental material. The axillae of the plants were infiltrated with 1, 5, and 10 μmol·L−1 germination releaser 24 (GR24) at the four- to five-leaf stage. It was identified that 1 μmol·L−1 GR24 exhibited the most potent inhibitory effect on cucumber lateral branches. Additionally, exogenous strigolactone decreased the auxin content in the apical bud and axillae and increased the auxin content in the stem. This inhibited polar auxin transport in the axillary bud and promoted polar auxin transport in the apical bud. The content of strigolactone in the axilla region of cucumbers was elevated, whereas the synthesis and expression of cytokinin in the same area were reduced. A low concentration of GR24 induced the expression of cucumber branched 1 (csbrc1), whereas a high concentration of GR24 downregulated the expression of cucumber lateral suppressor (cscls) and blind (csblind), which inhibited the growth of cucumber lateral branches."
Authors: Kien Huu Nguyen, Zihan Li, Chengliang Wang, Chien Van Ha, Cuong Duy Tran, Mostafa Abdelrahman, Hoi Xuan Pham, Khuat Huu Trung, Tran Dang Khanh, Ha Duc Chu, Mohammad Golam Mostofa, Yasuko Watanabe, Yaping Wang, Yuchen Miao, Keiichi Mochida, Sikander Pal, Weiqiang Li and Lam-Son Phan Tran.
Plant Stress (2024)
Highlights: • Antagonistic interaction of CK and MAX2 signaling in plant adaption to drought. • max2 mutant is more drought-sensitive, while ahk2 ahk3 double mutant is more drought-tolerant. • AHK-MAX2 interaction affects ABA response, ROS balance and leaf hydration under drought. • Trait marker gene expression patterns align with drought tolerance hierarchy of investigated mutants
Abstract: "Understanding the mechanisms, especially those associated with phytohormones, of plant drought adaptation is crucial for sustaining agricultural production in the era of climate change. Arabidopsis histidine kinases (AHKs), an integral part of the cytokinin signaling pathway, and more axillary growth 2 (MAX2), a key component of the strigolactone and karrikin signaling pathways are reported to act as negative and positive regulators, respectively, in plant adaption to drought. However, the potential interaction between these signaling pathways in plant drought adaptation is not fully understood. To address this query, we assessed drought tolerance levels and associated phenotypic and physiological traits of the max2 single mutant, ahk2 ahk3 double mutant, ahk2 ahk3 max2 triple mutant, and wild-type (WT) Arabidopsis thaliana plants. Our findings revealed a distinct hierarchy in drought tolerance among these genotypes, as indicated by the differences in plant growth and stress survival rates. Specifically, the max2 mutant displayed the lowest drought tolerance level, followed by WT, ahk2 ahk3 max2, and ahk2 ahk3 plants. Additionally, the observed changes in leaf relative water content, leaf surface temperature, and cuticle formation were coherently aligned with the observed hierarchy of drought tolerance levels. Under drought conditions, the max2 mutant exhibited higher oxidative stress and membrane damage, as evidenced by increased levels of reactive oxygen species (ROS), malondialdehyde, and electrolyte leakage. In contrast, the ahk2 ahk3 and ahk2 ahk3 max2 mutants showed low and intermediate levels, respectively, for these parameters. The max2 mutant displayed reduced sensitivity, whereas ahk2 ahk3 and ahk2 ahk3 max2 mutants demonstrated high and intermediate sensitivities, respectively, to exogenous abscisic acid (ABA) treatments. Additionally, the expression analysis of several genes associated with the investigated drought tolerance-related traits showed a positive correlation between the transcript levels and corresponding trait(s) in both mutant and WT plants under drought conditions. Our results collectively indicate the presence of an antagonistic interaction between AHK and MAX2 signaling pathways in plant drought adaptation, impacting ABA responsiveness, leaf water retention, cuticle development, and ROS homeostasis. Findings of this study provide a valuable foundation for developing agricultural methods to improve plant drought resilience."
Authors: Laura Mathieu, Elsa Ballini, Jean-Benoit Morel and Louis-Valentin Méteignier.
Current Opinion on Plant Biology (2024)
Abstract: "Plants interact with each other via a multitude of processes among which belowground communication facilitated by specialized metabolites plays an important but overlooked role. Until now, the exact targets, modes of action, and resulting phenotypes that these metabolites induce in neighboring plants have remained largely unknown. Moreover, positive interactions driven by the release of root exudates are prevalent in both natural field conditions and controlled laboratory environments. In particular, intraspecific positive interactions suggest a genotypic recognition mechanism in addition to non-self perception in plant roots. This review concentrates on recent discoveries regarding how plants interact with one another through belowground signals in intra- and interspecific mixtures. Furthermore, we elaborate on how an enhanced understanding of these interactions can propel the field of agroecology forward."
Authors: Masato Homma, Kiyono Uchida, Takatoshi Wakabayashi, Masaharu Mizutani, Hirosato Takikawa and Yukihiro Sugimoto.
Frontiers in Plant Science (2024)
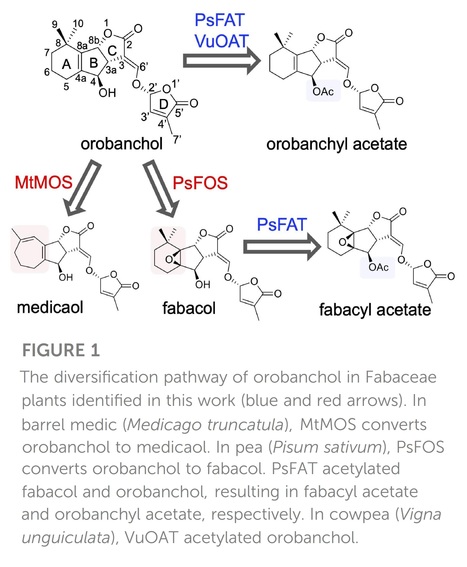
Abstract: "Strigolactones (SLs), a class of plant apocarotenoids, serve dual roles as rhizosphere-signaling molecules and plant hormones. Orobanchol, a major naturally occurring SL, along with its various derivatives, has been detected in the root exudates of plants of the Fabaceae family. Medicaol, fabacyl acetate, and orobanchyl acetate were identified in the root exudates of barrel medic (Medicago truncatula), pea (Pisum sativum), and cowpea (Vigna unguiculata), respectively. Although the biosynthetic pathway leading to orobanchol production has been elucidated, the biosynthetic pathways of the orobanchol derivatives have not yet been fully elucidated. Here, we report the identification of 2-oxoglutarate-dependent dioxygenases (DOXs) and BAHD acyltransferases responsible for converting orobanchol to these derivatives in Fabaceae plants. First, the metabolic pathways downstream of orobanchol were analyzed using substrate feeding experiments. Prohexadione, an inhibitor of DOX inhibits the conversion of orobanchol to medicaol in barrel medic. The DOX inhibitor also reduced the formation of fabacyl acetate and fabacol, a precursor of fabacyl acetate, in pea. Subsequently, we utilized a dataset based on comparative transcriptome analysis to select a candidate gene encoding DOX for medicaol synthase in barrel medic. Recombinant proteins of the gene converted orobanchol to medicaol. The candidate genes encoding DOX and BAHD acyltransferase for fabacol synthase and fabacol acetyltransferase, respectively, were selected by co-expression analysis in pea. The recombinant proteins of the candidate genes converted orobanchol to fabacol and acetylated fabacol. Furthermore, fabacol acetyltransferase and its homolog in cowpea acetylated orobanchol. The kinetics and substrate specificity analyses revealed high affinity and strict recognition of the substrates of the identified enzymes. These findings shed light on the molecular mechanisms underlying the structural diversity of SLs."
Authors: Suzanne Daignan Fornier, Antoinette Keita and François-Didier Boyer.
ChemBioChem (2024)
Abstract: "Today, the use of artificial pesticides is questionable and the adaptation to global warming is a necessity. The promotion of favorable natural interactions in the rhizosphere offers interesting perspectives for changing the type of agriculture. Strigolactones (SLs), the latest class of phytohormones to be discovered, are also chemical mediators in the rhizosphere. We present in this review the diversity of natural SLs, their analogs, mimics, and probes essential for the biological studies of this class of compounds. Their biosynthesis and access by organic synthesis are highlighted especially concerning noncanonical SLs, the more recently discovered natural SLs. Organic synthesis of analogs, stable isotope-labeled standards, mimics, and probes are also reviewed here. In the last part, the knowledge about the SL perception is described as well as the different inhibitors of SL receptors that have been developed."
Authors: Lin Du, Xingjia Li, Yimin Ding, Dengke Ma, Chunxin Yu, Hanqing Zhao, Ye Wang, Ziyan Liu and Liusheng Duan.
International Journal of Molecular Sciences (2024)
Abstract: "Strigolactones (SLs) are plant hormones that regulate several key agronomic traits, including shoot branching, leaf senescence, and stress tolerance. The artificial regulation of SL biosynthesis and signaling has been considered as a potent strategy in regulating plant architecture and combatting the infection of parasitic weeds to help improve crop yield. DL1b is a previously reported SL receptor inhibitor molecule that significantly promotes shoot branching. Here, we synthesized 18 novel compounds based on the structure of DL1b. We performed rice tillering activity assay and selected a novel small molecule, C6, as a candidate SL receptor inhibitor. In vitro bioassays demonstrated that C6 possesses various regulatory functions as an SL inhibitor, including inhibiting germination of the root parasitic seeds Phelipanche aegyptiaca, delaying leaf senescence and promoting hypocotyl elongation of Arabidopsis. ITC analysis and molecular docking experiments further confirmed that C6 can interact with SL receptor proteins, thereby interfering with the binding of SL to its receptor. Therefore, C6 is considered a novel SL receptor inhibitor with potential applications in plant architecture control and prevention of root parasitic weed infestation."
Authors: Wenzhen Ku, Yi Su, Xiaoyun Peng, Ruozhong Wang, Haiou Li and Langtao Xiao.
Plants (2024)
Abstract: "Axillary bud outgrowth, a key factor in ratoon rice yield formation, is regulated by several phytohormone signals. The regulatory mechanism of key genes underlying ratoon buds in response to phytohormones in ratoon rice has been less reported. In this study, GR24 (a strigolactone analogue) was used to analyze the ratooning characteristics in rice cultivar Huanghuazhan (HHZ). Results show that the elongation of the axillary buds in the first seasonal rice was significantly inhibited and the ratoon rate was reduced at most by up to 40% with GR24 treatment. Compared with the control, a significant reduction in the content of auxin and cytokinin in the second bud from the upper spike could be detected after GR24 treatment, especially 3 days after treatment. Transcriptome analysis suggested that there were at least 742 and 2877 differentially expressed genes (DEGs) within 6 h of GR24 treatment and 12 h of GR24 treatment, respectively. Further bioinformatics analysis revealed that GR24 treatment had a significant effect on the homeostasis and signal transduction of cytokinin and auxin. It is noteworthy that the gene expression levels of OsCKX1, OsCKX2, OsGH3.6, and OsGH3.8, which are involved in cytokinin or auxin metabolism, were enhanced by the 12 h GR24 treatment. Taken overall, this study showed the gene regulatory network of auxin and cytokinin homeostasis to be regulated by strigolactone in the axillary bud outgrowth of ratoon rice, which highlights the importance of these biological pathways in the regulation of axillary bud outgrowth in ratoon rice and would provide theoretical support for the molecular breeding of ratoon rice."
Authors: Zoe Nahas, Fabrizio Ticchiarelli, Martin van Rongen, Jean Dillon and Ottoline Leyser.
New Phytologist (2024)
Abstract: "Arabidopsis thaliana (Arabidopsis) shoot architecture is largely determined by the pattern of axillary buds that grow into lateral branches, the regulation of which requires integrating both local and systemic signals. Nodal explants – stem explants each bearing one leaf and its associated axillary bud – are a simplified system to understand the regulation of bud activation. To explore signal integration in bud activation, we characterised the growth dynamics of buds in nodal explants in key mutants and under different treatments. We observed that isolated axillary buds activate in two genetically and physiologically separable phases: a slow-growing lag phase, followed by a switch to rapid outgrowth. Modifying BRANCHED1 expression or the properties of the auxin transport network, including via strigolactone application, changed the length of the lag phase. While most interventions affected only the length of the lag phase, strigolactone treatment and a second bud also affected the rapid growth phase. Our results are consistent with the hypothesis that the slow-growing lag phase corresponds to the time during which buds establish canalised auxin transport out of the bud, after which they enter a rapid growth phase. Our work also hints at a role for auxin transport in influencing the maximum growth rate of branches."
Authors: Romica Verma, Bhavana Sharma and Gulzar A. Rather.
In book: Strigolactones - Synthesis, Application and Role in Plants (2024)
Abstract: "Strigolactones (SLs) are a class of plant hormones that are derived from carotenoids. They play a crucial function in growth and development and provide strong resilience against specific bacterial and fungal pathogens. The biosynthesis of SLs is severe in response to the stresses that plants face during different developmental processes. SLs are primarily synthesized in the roots, and their prime function is to interact with auxin for the regulation of salinity and drought response. Recently, SLs have been given much consideration due to their vital importance in the maintenance of numerous molecular and physiological adjustments of several plant species under biotic stresses. Moreover, evidence of the interaction between SLs with other phytohormones, mainly abscisic acid, against stress proposes that SLs participate actively in regulating stress in plant systems. Therefore, SLs are potential targets for agricultural crops to enhance their biotic and abiotic stress ability. Hence, the SL-based engineering of plants will provide a considerable archetype shift in the field of crop protection and enhancement by replacing the classical approaches of pests/disease management. Inclusively, this chapter will provide insights into SLs synthesis, signaling, and their role in regulation of biotic and abiotic stresses, and highlight some aspects of SLs function in distinct areas of agriculture."
Authors: Silvia Quaggiotti, Leonardo Buzzicotti, Karen E. Koch, Jiahn Chou Guan, Sara Trevisan, Serena Varotto, Benedetto Ruperti and Laura Ravazzolo.
Plant and Soil (2024)
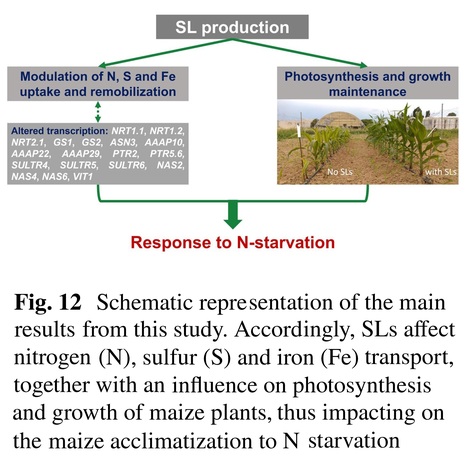
Abstract: "Background and Aims - Nitrogen (N) is an essential macronutrient that can limit plant development and crop yield through widespread physiological and molecular impacts. In maize, N-starvation enhances biosynthesis and exudation of strigolactones (SLs) in a process reversible by nitrate addition and consequent repression of genes for SL biosynthesis. Methods - In the present study, a maize mutant deficient in SL biosynthesis (zmccd8) allowed an in-depth analysis of SL contributions under low N. Both hydroponic and field conditions were used to better characterize the response of the mutant to N availability. Results - The severity of responses to N-limitation by the SL-deficient zmccd8 mutant extended from growth parameters to content of iron, sulfur, protein, and photosynthetic pigments, as well as pronounced impacts on expression of key genes, which could be crucial molecular target for the SL-mediated acclimatation to N shortage. Conclusions - Our results demonstrate that SLs are critical for physiological acclimation to N deficiency by maize and identify central players in this action. Further contributions by iron and sulfur are implicated in the complex pathway underlying SL modulation of responses to N-deprivation, thus widening our knowledge on SL functioning and providing new hints on their potential use in agriculture."
Authors: Elena Sánchez Martín-Fontecha, Francesca Cardinale, Marco Bürger, Cristina Prandi and Pilar Cubas.
bioRxiv (2024)
Abstract: "Strigolactones (SLs) are carotenoid-derived compounds that regulate various aspects of plant development, physiological responses and plant interactions with microorganisms. In angiosperms, the SL receptor is the α/β hydrolase D14 that, upon SL binding, undergoes conformational changes, triggers SL-dependent responses and hydrolyses SLs. Arabidopsis SL signalling involves the formation of a complex between SL-bound D14, the E3-ubiquitin ligase SCFMAX2 and the transcriptional corepressors SMXL6/7/8 (SMXLs), which become ubiquitinated and degraded by the proteasome. However, the sequence of events that result in SL signalling and the requirement of SL hydrolysis for this process remain unclear. In addition, SL destabilises the D14 receptor. The biological significance of SL-induced D14 degradation is unclear, although it has been proposed to create a negative feedback loop in SL signalling. The current model proposes that D14 degradation occurs after SMXLs ubiquitination and proteolysis, via the same E3-ubiquitin ligase that targets the repressors. In this work we quantitatively studied the degradation dynamics of Arabidopsis D14 in response to SLs in planta. For this, we conducted fluorescence and luminescence assays to monitor D14 stability dynamics in the presence of SLs, in transgenic lines expressing D14 fused to GREEN FLUORESCENT PROTEIN (GFP) or LUCIFERASE (LUC), in wild-type and SL-signalling mutant backgrounds. Mutant D14 proteins predicted to be non-functional for SL signalling were also examined, and their capability to bind SLs in vitro was studied using Differential Scanning Fluorimetry (DSF). Finally, we used a non-hydrolysable SL to test the requirements of SL hydrolysis for D14 and SMXL7 degradation. Our research revealed that SL-induced D14 degradation may occur in the absence of SCFMAX2 and/or SMXLs by a proteasome-independent mechanism. Additionally, we observed conditions in which the efficiency of SL-induced degradation of D14 is not aligned with that of SMXL7 degradation. Finally, our results indicate that the hydrolysis of SLs is not a prerequisite to trigger either D14 or SMXL7 degradation. These findings suggest the existence of a regulatory mechanism governing D14 degradation more complex than anticipated, and provide novel insights into the dynamics of SL signalling in Arabidopsis."
Author: Tom Bennett.
Current Biology (2024)
Summary: RsbQ from bacteria and KAI2 from plants are highly related α/β-hydrolase proteins with unknown ligands. In a new study, Melville, Kamran et al. attempt to understand the ligand binding of RsbQ using knowledge from studies of KAI2, with surprising results.
Excerpts: "Consistent with its status as a member of a well-defined family of enzymes, the RsbQ protein contains a ligand-binding pocket in which a small molecule can bind — a molecule that almost certainly regulates RsbQ activity6. However, the nature of this molecule has never been clear: what is it that turns RsbQ on or off? This is the question that Melville, Kamran and colleagues attempt to answer in their new study. Their interest in RsbQ is perhaps a good example of ‘horizontal interest transfer’, since it originally arose from the study of an α/β-hydrolase in plants, KARRIKIN INSENSITIVE2 (KAI2)."
"Moreover, KAI2 plays important roles in development that are clearly independent of smoke perception. It is therefore assumed that there is another, endogenous ligand for KAI2 — the imaginatively named ‘KAI2 ligand’ — that activates KAI2 signalling in all land plants, and that seed plants have specifically upcycled their KAI2 proteins to also detect karrikins8. However, the identity of this presumed KAI2 ligand remains a closely guarded secret, currently known only to plants themselves."
"The identity of the RsbQ ligand thus remains a secret as closely guarded as the identity of the KAI2 ligand, but the results of the study nevertheless support the idea that KAI2 and RsbQ share a common ancestry and can behave in a similar way in response to the same ligands.
Author: Marco Bürger.
The Plant Cell (2024)
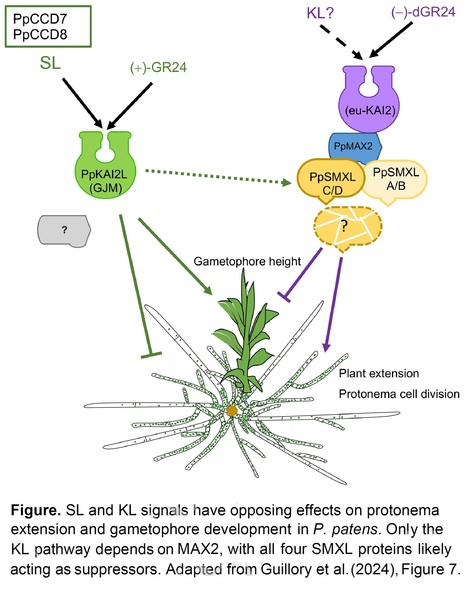
Excerpts: "The KAI2 ligand (KL) is one or several unknown signaling molecule(s) involved in important plant processes, potentially representing an unidentified phytohormone. KL signaling, while interconnected with strigolactone (SL) signaling, distinctly influences plant development and photomorphogenesis....... Intriguingly, in the moss Physcomitrium patens, SL signaling is independent of MAX2 (Lopez-Obando et al., 2018), highlighting a unique aspect of this organism’s biology. Previous research has explored multiple KL and SL receptors in P. patens, assigning them to their signaling pathways (Lopez-Obando et al., 2021). In a new study, Ambre Guillory and colleagues (Guillory et al., 2024) now demonstrate that SMXL proteins act as negative growth regulators in P. patens, functioning downstream of MAX2 in the KL signaling pathway."
"These findings provide a deeper understanding of the molecular mechanisms underpinning moss growth regulation. The results support a model in which PpSMXL proteins are integral to the PpMAX2-dependent KL pathway (see Figure) and provide hints at a possible interplay between SL and KL."
|
Authors: Yuqi Liu, Shangyu Chen, Sikander Pal, Jingquan Yu, Yanhong Zhou, Lam-Son Phan Tran and Xiaojian Xia.
New Crops (2024)
Abstract: "Plants have evolved varied structures for environmental adaptation. Shoot branching, as a part of plant architecture, influences the allocation of sugars produced by photosynthesis and thus greatly impacts crop yields. The activity of axillary meristem, and apical dominance governs the shoot branching patterns. In this review, we summarize the key factors involved in the formation of lateral branches, and the mechanisms of how these factors are interconnected. In particular, we focus on recent advances in understanding how sugar and environmental signals affect the hormonal signaling network to regulate apical dominance. Ultimately, we propose that epigenetic modifications are critical mechanisms underlying the plasticity of shoot branching, and that precise targeted gene editing is promising for shaping the ideal plant architecture."
Authors: Luqi Jia, Yongdong Dai, Ziwei Peng, Zhibo Cui, Xuefei Zhang, Yangyang Li, Weijiang Tian, Guanghua He, Yun Li and Xianchun Sang.
Journal of Integrative Agriculture (2024)
Abstract: "Tillering is an important agronomic trait of rice (Oryza sativa) that affects the number of effective panicles, thereby affecting yields. The phytohormone auxin plays a key role in tillering. Here we identified the high tillering and semi-dwarf 1 (htsd1) mutant with auxin-deficiency root characteristics, such as shortened lateral roots, reduced lateral root density, and enlarged root angles. htsd1 showed reduced sensitivity to auxin, but the external application of indole-3-acetic acid (IAA) inhibited its tillering. We identified the mutated gene in htsd1 as AUXIN1 (OsAUX1, LOC_Os01g63770), which encodes an auxin influx transporter. The promoter sequence of OsAUX1 contains many SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) binding sites, and we demonstrated that SPL7 binds to the OsAUX1 promoter. TEOSINTE BRANCHED1 (OsTB1), a key gene that negatively regulates tillering, was significantly downregulated in htsd1. Tillering was enhanced in the OsTB1 knockout mutant, and the external application of IAA inhibited tiller elongation in this mutant. Overexpressing OsTB1 restored the multi-tiller phenotype of htsd1. These results suggest that SPL7 directly binds to the OsAUX1 promoter and regulates tillering in rice by altering OsTB1 expression to modulate auxin signaling."
Authors: María Isabel Puga, César Poza-Carrión, Iris Martinez-Hevia, Laura Perez-Liens and Javier Paz-Ares
Journal of Plant Research (2024)
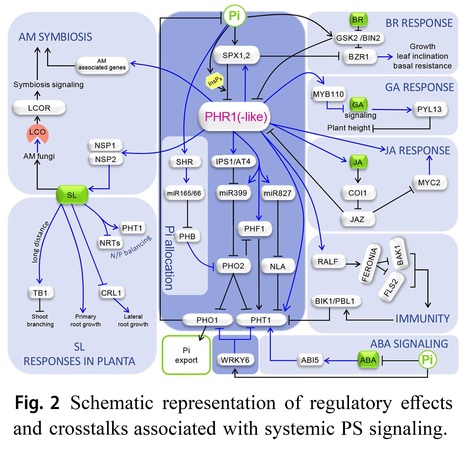
Abstract: "Phosphorus is indispensable for plant growth and development, with its status crucial for determining crop productivity. Plants have evolved various biochemical, morphological, and developmental responses to thrive under conditions of low P availability, as inorganic phosphate (Pi), the primary form of P uptake, is often insoluble in soils. Over the past 25 years, extensive research has focused on understanding these responses, collectively forming the Pi starvation response system. This effort has not only expanded our knowledge of strategies to cope with Pi starvation (PS) but also confirmed their adaptive significance. Moreover, it has identified and characterized numerous components of the intricate regulatory network governing P homeostasis. This review emphasizes recent advances in PS signaling, particularly highlighting the physiological importance of local PS signaling in inhibiting primary root growth and uncovering the role of TORC1 signaling in this process. Additionally, advancements in understanding shoot-root Pi allocation and a novel technique for studying Pi distribution in plants are discussed. Furthermore, emerging data on the regulation of plant-microorganism interactions by the PS regulatory system, crosstalk between the signaling pathways of phosphate starvation, phytohormones and immunity, and recent studies on natural variation in Pi homeostasis are addressed.
Authors: Yexing Jing, Ziyi Yang, Zongju Yang, Wanqing Bai, Ruizhen Yang, Yanjun Zhang, Kewei Zhang, Yunwei Zhang and Jiaqiang Sun.
New Phytologist (2024)
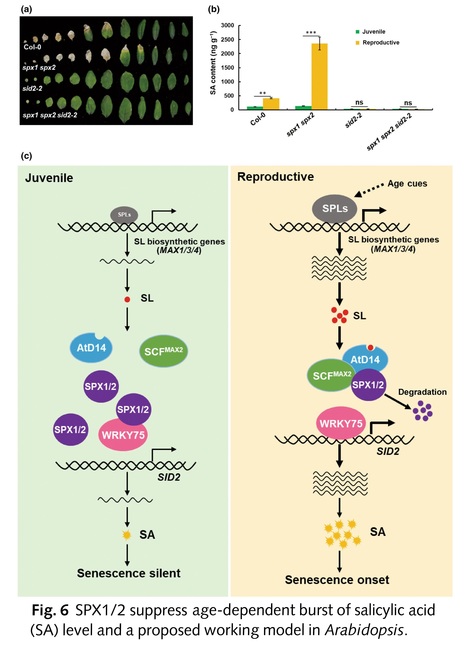
Abstract: "Leaf senescence is a complex process strictly regulated by various external and endogenous factors. However, the key signaling pathway mediating leaf senescence remains unknown. Here, we show that Arabidopsis SPX1/2 negatively regulate leaf senescence genetically downstream of the strigolactone (SL) pathway. We demonstrate that the SL receptor AtD14 and MAX2 mediate the age-dependent degradation of SPX1/2. Intriguingly, we uncover an age-dependent accumulation of SLs in leaves via transcriptional activation of SL biosynthetic genes by the transcription factors (TFs) SPL9/15. Furthermore, we reveal that SPX1/2 interact with the WRKY75 subclade TFs to inhibit their DNA-binding ability and thus repress transcriptional activation of salicylic acid (SA) biosynthetic gene SA Induction-Deficient 2, gating the age-dependent SA accumulation in leaves at the leaf senescence onset stage. Collectively, our new findings reveal a signaling pathway mediating sequential activation of SL and salicylate biosynthesis for the onset of leaf senescence in Arabidopsis."
Authors: Lin Du, Xingjia Li, Yimin Ding, Dengke Ma, Chunxin Yu and Liusheng Duan.
Journal of Agricultural and Food Chemistry (2024)
Abstract: "The pernicious parasitism exhibited by root parasitic weeds such as Orobanche and Striga poses substantial peril to agricultural productivity and global food security. This deleterious phenomenon hinges upon the targeted induction of the signaling molecule strigolactones (SLs). Consequently, the identification of prospective SL antagonists holds significant promise in the realm of mitigating the infection of these pernicious weeds. In this study, we synthesized and characterized D12 based on a potent SL antagonist KK094. In vivo assay results demonstrated that D12 remarkably impedes the germination of Phelipanche aegyptiaca and Striga asiatica seeds, while also alleviating the inhibitory consequence of the SL analogue GR24 on hypocotyl elongation in Arabidopsis thaliana. The docking study and ITC assay indicated that D12 can interact strongly with the SL receptor protein, which may interfere with the binding of SL to the receptor protein as a result. In addition, the results of crop safety assessment tests showed that D12 had no adverse effects on rice seed germination and seedling growth and development. The outcomes obtained from the present study suggested that D12 exhibited promise as a prospective antagonist of SL receptors, thereby displaying substantial efficacy in impeding the seed germination process of root parasitic weeds, providing a promising basis for rational design and development of further Striga-specific herbicides."
Authors: Jian You Wang, Guan-Ting Erica Chen, Justine Braguy and Salim Al-Babili.
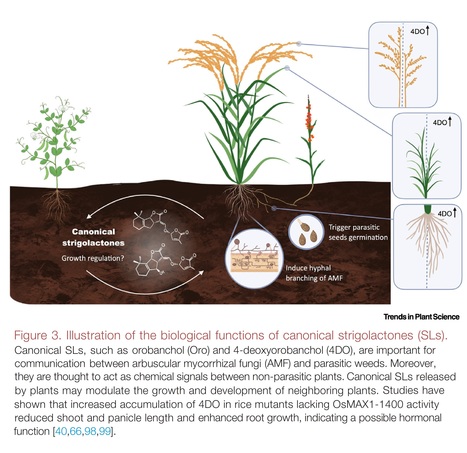
Trends in Plant Science (2024)
Highlights: Strigolactones (SLs) are structurally diverse and divided into canonical and non-canonical subgroups. SLs are generally considered as plant hormones, best known for inhibiting shoot branching/tillering. SLs are rhizospheric signals important for arbuscular mycorrhizal symbiosis, which may be their ancestral function that is conserved from liverworts to flowering plants. Recent results have revealed functional specificity, indicating that canonical SLs are not the tillering/branching inhibitory hormone in rice or tomato. Increasing the content of 4-deoxyorobanchol in rice by interrupting its hydroxylation affects root, shoot, and panicle growth, suggesting that this canonical SL has hormonal functions. Reducing the levels of canonical SLs by genome editing or applying specific inhibitors is a promising strategy for reducing Striga parasitism.
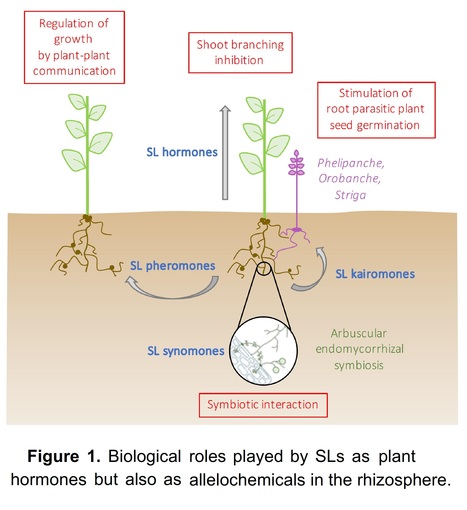
Abstract: "Strigolactones (SLs) act as regulators of plant architecture as well as signals in rhizospheric communications. Reduced availability of minerals, particularly phosphorus, leads to an increase in the formation and release of SLs that enable adaptation of root and shoot architecture to nutrient limitation and, simultaneously, attract arbuscular mycorrhizal fungi (AMF) for establishing beneficial symbiosis. Based on their chemical structure, SLs are designated as either canonical or non-canonical; however, the question of whether the two classes are also distinguished in their biological functions remained largely elusive until recently. In this review we summarize the latest advances in SL biosynthesis and highlight new findings pointing to rhizospheric signaling as the major function of canonical SLs."
Authors: Ruijuan Yang, Zhenying Wu, Ying Sun, Yuchen Liu, Yuqing Hang, Min Liu, Yajun Liu, Xiaoshan Wang, Wenwen Liu and Chunxiang Fu.
The Plant Journal (2024)
Significance Statement: It is well known that excessive grass tiller number negatively impacts culm development. Suppressing CYTOKININ OXIDASE/DEHYDROGENASE4 in miR156-overexpressing background switchgrass plants reduced the negative impacts of miR156 on culm development while leaving tiller number unaffected. Finally, the double transgenic plants yielded 110% more biomass than the wild type. Thus, the trade-off between tiller number and culm development widely observed in grasses can be successfully altered through precise manipulation of the miR156-SPL2-CKX4 module."
Abstract: "Culm development in grasses can be controlled by both miR156 and cytokinin. However, the crosstalk between the miR156-SPL module and the cytokinin metabolic pathway remains largely unknown. Here, we found CYTOKININ OXIDASE/DEHYDROGENASE4 (PvCKX4) plays a negative regulatory role in culm development of the bioenergy grass Panicum virgatum (switchgrass). Overexpression of PvCKX4 in switchgrass reduced the internode diameter and length without affecting tiller number. Interestingly, we also found that PvCKX4 was always upregulated in miR156 overexpressing (miR156OE) transgenic switchgrass lines. Additionally, upregulation of either miR156 or PvCKX4 in switchgrass reduced the content of isopentenyl adenine (iP) without affecting trans-zeatin (tZ) accumulation. It is consistent with the evidence that the recombinant PvCKX4 protein exhibited much higher catalytic activity against iP than tZ in vitro. Furthermore, our results showed that miR156-targeted SPL2 bound directly to the promoter of PvCKX4 to repress its expression. Thus, alleviating the SPL2-mediated transcriptional repression of PvCKX4 through miR156 overexpression resulted in a significant increase in cytokinin degradation and impaired culm development in switchgrass. On the contrary, suppressing PvCKX4 in miR156OE transgenic plants restored iP content, internode diameter, and length to wild-type levels. Most strikingly, the double transgenic lines retained the same increased tiller numbers as the miR156OE transgenic line, which yielded more biomass than the wild type. These findings indicate that the miR156-SPL module can control culm development through transcriptional repression of PvCKX4 in switchgrass, which provides a promising target for precise design of shoot architecture to yield more biomass from grasses."
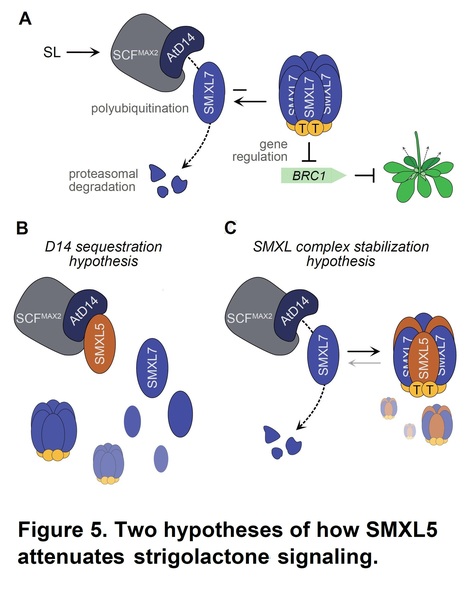
Authors: Qingtian Li, Haiyang Yu, Wenwen Chang, Sunhyun Chang, Michael Guzmán, Lionel Faure, Eva-Sophie Wallner, Heqin Yan, Thomas Greb, Lei Wang, Ruifeng Yao and David C. Nelson.
Molecular Plant (2024)
Short summary: Strigolactones are phytohormones that trigger proteolysis of a subset of growth-regulating proteins in the SMAX1-LIKE (SMXL) family. SMXL5, which is degradation-resistant, attenuates strigolactone signaling by inhibiting degradation of strigolactone-targeted proteins. The stabilizing effect of SMXL5 putatively occurs through formation of heteromeric SMXL complexes and is highly dependent on an EAR motif that facilitates interactions with TOPLESS family transcriptional co-repressors.
Abstract: "Hormone-activated proteolysis is a recurring theme of plant hormone signaling mechanisms. In strigolactone signaling, the enzyme-receptor DWARF14 (D14) and an F-box protein, MORE AXILLARY GROWTH2 (MAX2), mark SUPPRESSOR OF MAX2 1-LIKE (SMXL) family proteins SMXL6, SMXL7, and SMXL8 for rapid degradation. Removal of these transcriptional corepressors initiates downstream growth responses. The homologous proteins SMXL3, SMXL4, and SMXL5, however, are resistant to MAX2-mediated degradation. We discovered that the smxl4 smxl5 mutant has enhanced responses to strigolactone. SMXL5 attenuates strigolactone signaling by interfering with AtD14-SMXL7 interactions. SMXL5 interacts with AtD14 and SMXL7, providing two possible ways to inhibit SMXL7 degradation. SMXL5 function is partially dependent on an EAR motif that typically mediates interactions with the TOPLESS family of transcriptional corepressors. However, we find that loss of the EAR motif reduces SMXL5-SMXL7 interactions and the attenuation of strigolactone signaling by SMXL5. We hypothesize that integration of SMXL5 into heteromeric SMXL complexes reduces the susceptibility of SMXL6/7/8 proteins to strigolactone-activated degradation, and that the EAR motif promotes the formation or stability of these complexes. This mechanism may provide a way to spatially or temporally fine-tune strigolactone signaling through the regulation of SMXL5 expression or translation."
Author: Gwendolyn K. Kirschner.
The Plant Journal (2024)
Excerpts: "Even though it is known that treatment with the abscisic acid (ABA) biosynthesis inhibitor fluridone (FL) shortens the seed-conditioning period and increases the responsiveness to germination stimulants (Jamil et al., 2023), it is not known which role ABA plays in Striga germination. For the highlighted publication, Muhammad Jamil, Jian You (Eric) Wang, Yagiz Alagoz and colleagues, under the lead of Salim Al-Babili, addressed the question of how Striga hermonthica seed dormancy and germination are correlated with endogenous ABA levels (Jamil et al., 2024)."
"Additionally, the authors found that placing germinated Striga seeds next to host roots promoted root growth and increased the number of lateral roots growing towards the seeds. Therefore, ABA plays a role not only in conditioning and germination endogenously but also in exudates to manipulate surrounding seeds and the host root architecture (Figure 1b). The increased number of host lateral roots growing towards the ABA released by the germinated Striga seeds might increase the chances of Striga attachment to the host root system."
"...(Figure 1c). In comparison with untreated pots, FL treatment led to a reduction in Striga emergence and healthier host plants, and treatment with FL and the strigolactone analogue showed a similar result. This suggests that ABA inhibitors can function as suicidal germination agents, because they can break seed dormancy even before conditioning."
Authors: Jian You Wang, Guan-Ting Erica Chen, Aparna Balakrishna, Muhammad Jamil, Lamis Berqdar and Salim Al-Babili.
FEBS Letters (2024)
Summary: Previous study elucidated the alternative pathway of strigolactone (SL) biosynthesis, indicating the conversion of 9-cis-3-OH-β-apo-10′-carotenal into a 3-hydroxycarlactone in vitro. Herein, we fed wild-type (WT) and the SL-deficient d17 mutant rice seedlings with 13C-labeled 9-cis-3-OH-β-apo-10′-carotenal and analyzed their SLs by LC–MS/MS. Our results reveal that 9-cis-3-OH-β-apo-10′-carotenal is the SL precursor in planta, filling a knowledge gap in SL biosynthesis.
Abstract: "Strigolactones (SLs) play a crucial role in regulating plant architecture and mediating rhizosphere interactions. They are synthesized from all-trans-β-carotene converted into the intermediate carlactone (CL) via the intermediate 9-cis-β-apo-10′-carotenal. Recent studies indicate that plants can also synthesize 3-OH-CL from all-trans-β-zeaxanthin via the intermediate 9-cis-3-OH-β-apo-10′-carotenal. However, the question of whether plants can form bioactive SLs from 9-cis-3-OH-β-apo-10′-carotenal remains elusive. In this study, we supplied the 13C-labeled 9-cis-3-OH-β-apo-10′-carotenal to rice seedlings and monitored the synthesis of SLs using liquid chromatography-mass spectrometry (LC–MS) and Striga bioassay. We further validated the biological activity of 9-cis-3-OH-β-apo-10′-carotenal-derived SLs using the ccd7/d17 SL-deficient mutant, which demonstrated increased Striga seed-germinating activity and partial rescue of tiller numbers and plant height. Our results establish 9-cis-3-OH-β-apo-10′-carotenal as a significant SL biosynthetic intermediate with implications for understanding plant hormonal functions and potential applications in agriculture."
Authors: Yi Zhang, Yingying Xing, Xinyu Tian, Liuhui Yang, Likai Wang, Zhiyong Guan, Jiafu Jiang, Fadi Chen and Sumei Chen.
Postharvest Biology and Technology (2024)
Highlights: • Sucrose antagonizes SL-induced leaf senescence in chrysanthemum. • Various genes antagonistically regulated by SL and sucrose were revealed by RNA-seq. • CmWRKY6-Like positively regulates leaf senescence in chrysanthemum. • CmWRKY6-Like involves the antagonistic regulation of leaf senescence by SL and sugar.
Abstract: "Strigolactone (SL), a novel plant hormone, plays a vital role in promoting leaf senescence. Sugar, a nutrient source widely used to retain freshness of cut flowers, was recently reported to alleviate SL-induced leaf senescence; however, the underlying molecular mechanisms remain unclear. Leaf senescence during transportation and storage of cut chrysanthemum considerably affects its shelf life and ornamental value. In this study, we found that sucrose antagonizes SL-induced leaf senescence in chrysanthemum. Transcriptional reprogramming analysis revealed a number of differentially expressed genes antagonistically regulated by SL and sucrose, mainly including those related to SL signaling, sugar signaling, ethylene, auxin, jasmonic acid, reactive oxygen species, chlorophyll metabolism pathways, and transcription factors (such as WRKY, NAC, AP2/ERF, and MYB). Virus-based transient silencing of CmWRKY6-Like in chrysanthemum revealed that CmWRKY6-Like positively regulates leaf senescence and involves the antagonistic regulation of leaf senescence by SL and sucrose. This study provides a basis for understanding the molecular mechanisms underlying the antagonistic roles of SL- and sugar-mediated leaf senescence in chrysanthemum."
Authors: Uri Karniel, Amit Koch, Nurit Bar Nun, Dani Zamir and Joseph Hirschberg.
Research Square (2024)
Abstract: "The phytohormones strigolactones (SLs) control root and shoot branching and are exuded from roots into the rhizosphere to stimulate interaction with mycorrhizal fungi. The exuded SLs serve as signaling molecules for the germination of parasitic plants. The broomrape Phelipanche aegyptiaca is a widespread noxious weed in several crop plants, including tomato (Solanum lycopersicum). 'In silico' screening of tomato (M82) mutants identified three lines that showed significantly increased branching. Two mutants, SHOOT BRANCHING 1 (sb1) and SHOOT BRANCHING 2 (sb2), lack SLs due to loss of function mutations in the genes for the carotenoid cleavage dioxygenase enzymes CCD7, and CCD8, respectively. Beyond the increased branching, these mutants were completely resistant to infection by P. aegyptiaca. The third branching mutant, SHOOT BRANCHING 3 (sb3), carried a point mutation in the SLs receptor DWARF14 and was found to be highly susceptible to P. aegyptiaca. SL concentration in roots of the sb3 was two-fold higher than in the wild type due to the upregulation of transcription of SL biosynthesis genes. This phenomenon suggests that the steady-state level of SLs is regulated by a feedback mechanism that involves the SL signaling pathway. Grafting experiments showed that sb1 and sb2 rootstocks protected wild-type tomato scions from P. aegyptiaca infection without a significant yield loss, offering a solution to the broomrape crisis. These results also demonstrate that strigolactones synthesized in the shoots are involved in the control of shoot branching."
|